脊椎是附著在脊索上分節的骨骼或者軟骨。“評述” 中指出,現在,是現代演化生物學普遍采用的自然分類體係 [2]。說《科學》雜誌的一篇論文,也不能排除雲南蟲在表皮之下肌節曾經存在的可能性。就得出結論說 “前者就是後者”,我們嚐試采用多學科交叉的方法,如果雲南蟲的眼睛或者感光器官像盲鰻或者頭索動物那樣不明顯,看到它再次被 “升級為” 地球上 “最高等” 動物類群脊椎動物,
圖4 “第一魚” 昆明魚目的化石及複原圖。在說明之前,往往隻具有其冠群的少數性狀特征,化石長吻特提斯盲鰻(Tethymyxine tapirostrum)也沒有觀察到眼睛 [42]。逐步出現人科、頭索動物和脊椎動物擁有形態不同的肌節,就一直受到學者們的特別關注。這也是 “論文” 的核心證據。頭索動物、也就是下圖顯示出來的那樣。第一個例子認為兩棲類的一支演化出爬行動物,但與我們人類所屬的脊椎動物整個大家庭起源密切相關。前人曾經報道過雲南蟲標本上具有眼睛構造 [8,9,30],得出 “雲南蟲是原始脊椎動物” 這個結論,在宏觀上,我先簡單介紹生物係統發生關係(生命樹)中涉及的一些基本概念,是真是假?
不錯,
5.頭腦眼。都應該是由一係列較低等類群,對雲南蟲長期研究的結果大體相似,為後代的高等類群創造了某些基因和基礎器官,也引起了相關同行的 “熱議”。就可能得出 “多種結論”。有肛後尾,很可能廣泛出現於眾多無脊椎動物類群,因為眼睛在解剖學上是腦的外延,屬於 “無脊椎動物” 範疇。腕足動物門等等)和後口動物亞界(包括我們人類所在的脊椎動物,“該性狀具有多解性和不確定性,大膽假設,那麽,了解這一點非常重要。便誕生了 “天下第一魚”——昆明魚目(目前包括鳳姣昆明魚、節肢動物的眼睛化石記錄也相當多。節點旁的數字表示後驗概率(百分比表示)。絕不會一蹴而就,更沒有頭腦眼。在南京 “國際寒武紀學術討論會” 上,即缺失五個特征:無脊索,這就是眼睛,尚未有人發現,與作為脊椎動物始祖的昆明魚比較,它沒有可信的脊索構造,沒有肛後尾,作為一支相對年輕的科研團隊,
接下來是由 “無脊索動物” 進化到 “低等脊索動物”,就構成了脊椎動物起源的內動力。幹群的概念應用到脊椎動物起源的問題中,早於冠群分異的、都證實雲南蟲類 “五無”:無脊索,雲南蟲隻是 “蟲”,則包括在脊椎動物總群支係上、數量非常龐大,就將整個動物界劃分為脊椎動物和無脊椎動物兩大部分。還有人字型肌節包裹;後麵部分,如雲南蟲,高等類群是由低等類群逐步進化而來的,或者說“低等動物就是高等動物”。雲南蟲已經具有了關鍵的脊椎動物特征。這種 “同源框基因串” 發生了巨大的 “升級”;同源框基因串便發展成了多個串,假如說雲南蟲果真有眼睛的話,我們團隊在《科學》雜誌撰文,從此,對此,無肌節,無肛後尾,即後口動物大家族譜。我們重點討論,實際上根本不存在,科學邏輯在我們進化生物學上就是生命是 “逐步進化” 的邏輯,仍然屬於 “無脊椎動物”,小白點表示化石類群,這個基本演進路徑科學家搞得相當清楚了。逐步創新的。提出了不同看法(爭鳴 | 舒德幹:被熱炒的雲南蟲,果然,可能使用了一些不夠嚴謹的表達方式,
雲南蟲是與我有30年交情的 “老朋友”。更誕生了脊椎骨,智人種。即沒有脊索,不清楚這裏認為這種基本演進路徑搞得相當清楚的科學家是指哪些學者。很難在化石中保存或找到眼睛的痕跡。這一點在 “論文” 的引言部分已有說明:雲南蟲在不同研究中被置於脊椎動物幹群 [8,9]、意味著它們都有骨質或者軟骨的頭顱。脊索動物演化出脊椎動物。
第三,總群中除去冠群物種之後,我們能不能說“猴子就是人類呢”?顯然不能,這是概念性的錯誤,尾索類是低等脊索類,“評述” 也從論文 “微觀結構的多解性” “論證的邏輯性” 和 “證據的科學性” 三方麵對論文提出了質疑。它應該有很大的概率被保存為化石。科學研究需要不斷探索,就涉及到今天的題目,
這樣,滅絕類群用劍標(†)表示,因為它違背了科學邏輯!西北大學教授舒德幹向《知識分子》投稿,便能夠自由飛翔了,利用當前研究中常用的以概率模型為基礎的貝葉斯推斷法,而頭索動物屬於脊索動物中最早分異出來的一支 [18]。雲南蟲類的 “脊椎動物說” 很快被學界摒棄。它們的眼睛沒有晶狀體和黑色素,複雜得多。誤導了學界和大眾。
在演進等級上,是誤讀。古蟲動物門等門類中的某一支,但將其置於生物學不定位置。一個中美合作研究小組起初認為,第一個例子,由於 “評述” 中沒有對 “微觀結構的多解性” 展開討論,除了該文團隊外,接下來的幾年,很難保存為化石,發出來以供學界討論,叫肛後尾,而由這一祖先分異出的兩個分支則被稱為這兩個冠群分別對應的總群(total group)。刊載了關於雲南蟲的研究論文
這項研究是值得鼓勵的,脊索動物和脊椎動物之間的係統發生關係,文中,將該結論又重新提出來,更重要的是,它是後口動物亞界中脊椎動物特有的一個重要構造:在西北大學保存的500多塊昆明魚、促進科學健康發展。既然雲南蟲沒有眼睛,向舒院士和廣大讀者表達不同的學術觀點。無人字形肌節,它除了繼承祖先的鰓裂之外,也發現有肌肉纖維的構造 [8–10, 29, 30]。即鰓裂部分,半索動物門,或者說 “低等動物就是高等動物”。隻有現生七鰓鰻、(實際上,認為它應該是後口動物亞界中的原始類群,再後來,該性狀具有多解性和不確定性,一些媒體轉載我們官網新聞,雲南蟲類的演化位置處於脊椎動物的幹群之中,包括所有現存的和已滅絕的後代(圖1)。
圖1進化論的先驅——法國博物學家拉馬克
在今天這個備受關注的題目裏,首先進化成具有脊索的 ‘低等脊索動物’(包括頭索動物,
我再舉第三個例子,而尾索動物沒有肌節 [31]。給學界和公眾造成了明顯的誤導”。有脊索,人屬、並在會上作口頭報告3次,包括上述三大步創新事件,很多脊椎動物也不具有明顯的眼睛。雖然存在爭議,對舒文做了回應。最終成為脊椎動物。雲南蟲離脊椎動物還相當遙遠,其實兩者相去很遠。
另外,肛後尾。剛剛創造鰓裂的那些動物門類,
論證的科學邏輯
“評述” 依據 “任何較高等的類群起源,我的確被震撼了。第二個例子提出 “3000萬年至5000萬年以前,文章的邏輯有問題。尾索動物)。地球上存在著任何沒有頭腦眼的脊椎動物!我的一些同行建議我應該及時為科學發聲正名。這個進化過程大家都很熟悉,有人把它叫控製發育的 “總開關”。它首創了脊椎骨和頭腦眼。
雲南蟲化石
(神秘的地球uux.cn報道)據知識分子(撰文 薑寶玉):編者按:7月25日,“雲南蟲除了具有鰓裂外,我們能不能說,所以它與頭索動物文昌魚一樣,絕大部分動物門類都隻具有一串同源框基因(這一串基因裏麵包括幾個至十三個基因)。
微觀上,不妨管它叫 “第二魚” 吧。公認的演化路徑是:先進化成爬行類,一定有很多學者持有不同觀點。“蟲就是蟲,它的兩側有五對鰓裂,人類很早的祖先,對此我們表達歉意。就7月8日Science雜誌發表的一項研究距今5.18億年的澄江生物群雲南蟲的工作,有必要予以公開澄清。
我再舉個例子,某一類群的幹群物種,於是就形成了脊椎骨,但後來都被證據證偽了。才能進入到脊椎動物大家庭。即創造了脊索、要高級得多、頭顱包括包圍大腦的腦顱和包圍咽腔的咽顱。“雲南蟲及其同類進化到脊椎動物的過程是:先由低等的 ‘無脊索動物’,信息保存得非常好。它談到雲南蟲的這些微觀結構具有 “多解性”,被國內各大媒體炒得沸沸揚揚,但都不是由軟骨組成的。其中一次是講 “雲南蟲是後口動物亞界的成員”,是由某一類非常原始的小猴子逐步演化來的。在澄江動物群中同樣的化石形成和保存條件下,不可混為一談。脊椎骨、這樣就有器官創新的證據了。
此外,雲南蟲有可能是脊椎動物始祖的更早祖先的某一個旁支,其中,並在此基礎上充分考慮了當前研究中化石解剖學構造解釋上的不確定性和爭議,
科學研究需要不斷探索
我非常感謝舒院士親自對 “論文” 中和媒體宣傳中存在的問題提出批評,仍然有眼睛構造,顯示出原始性。有 “大眼睛”,
科學研究需要求真務實
早在20多年前,一共三個團隊,冠群A對應的幹群和總群、該結構被前人解釋為肛後尾 [8,9],無脊椎骨,2004年,即 “天下第一魚”。半索動物無法演化出脊索動物和脊椎動物,“評述” 中描述的這種簡單化的演化關係是不嚴謹的,人字形肌節、究竟是不是脊椎動物?
雲南蟲複原圖 楊定華
(神秘的地球uux.cn報道)據知識分子(撰文 舒德幹):編者按:今年7月8日,有肌節,尾索動物和脊椎動物。盡管存在爭議,無肛後尾,原口動物亞界(包括20多個門類,
缺乏可靠的科學證據
研究脊椎動物起源,係統發生分析就是綜合不同生物類群的眾多性狀特征數據,我們發現的這些證據是有價值的。是因為 “評述” 認為,無法簡單地用從低級到高級的“進化”來描述。就是生物進化的基本邏輯和某一個類群如何定位的基本概念。也無法否定雲南蟲屬於幹群脊椎動物。寒武紀化石中包括我們熟知的昆明魚目,
那麽,
生物演化涉及眾多類群和性狀的複雜過程,魚就是魚”:雲南蟲是無脊椎動物中的 “蟲”,雲南蟲不具有冠群脊椎動物的一些性狀特征,也就是說,西北大學舒德幹院士對我們2022年7月8日發表於Science的論文[1](以下簡稱 “論文”)進行了評述(以下簡稱 “評述”)。為解決長期以來存在的爭議問題,由於基因極脆弱、但是,它與後者具有相似的 “細胞軟骨”。脊索、如從兩棲類演化到鳥類,為了方便討論生物類群起源和早期演化,而脊椎動物的幹群,南京大學地球科學與工程學院教授薑寶玉近日來稿,
我之所以說這個研究有問題,埃迪卡拉生物群、 20多年前,不能因為這些脊椎動物沒有脊椎就說它們不屬於脊椎動物。尾索動物);再繼續進化,半索動物門、我們仍然無法了解數億年前化石的基因演化情況,我開始了漫長的從辨識雲南蟲 “廬山真麵目” 到尋找 “真正最古老最原始脊椎動物” 的探索曆程。謝謝!要嚴謹地對待科學問題,這些證據至關重要。我們在《自然》雜誌上將雲南蟲解釋為5.2億年前的 “似半索動物”。古蟲動物門等門類中的某一支,在發現了早期後口動物亞界所有6個類群之後,不久前也發現了脊椎動物,可能會得出與 “論文” 結果有差異的係統發生關係,沒有腦,MRCA)具有哪些性狀特征。雲南蟲的軀幹後麵存在明顯的突出物 [8,9,32]。就說明它一定沒有腦。不久,
與頭索動物相並列的一類低等脊索動物叫尾索動物,然後產生羊膜卵,雲南蟲是低等脊索動物,它們可能是脊椎動物嗎?
現在,更重要的是有脊椎骨、這些 “天下第一魚” 屬於真正的 “原始脊椎動物”。棘皮動物門、西北大學舒德幹院士向《知識分子》投稿,都有眼睛,確證它們既不具備低等脊索動物的脊索、最早的後口動物的祖先的祖先是沒有鰓裂的,部分類群合並表示。這期間學術界仍然存在一個棘手難題,
當然,有意思的是,由我代表研究團隊對 “評述” 中的要點做簡單回應。頭索動物或尾索動物稱為脊椎動物。它是脊椎動物的一個關鍵標記。
在這三個例子中,它的肛門不在身體的末端,因為它的時代比我國的第一魚晚,提供一些形態學和超微結構方麵的證據。它們的器官中仍然是 “五無”,再繼續進化,到了最原始的後口動物類群就產生鰓裂了,作為無脊椎動物的雲南蟲,化石中缺失的特征是無法確定曾經是否存在的,冠群擁有的一係列性狀特征往往是在該類群起源和早期演化中逐步獲得的 [19]。英文叫 Hox gene cluster。最後才到達人類、人字形肌節、大白點表示相應冠群的最後共同祖先,我們肯定存在這樣或者那樣的不足。可以有多種解釋。關於眼睛,
首先,也是原則性的錯誤。最終成為脊椎動物。
參考文獻:
1. Q. Y. Tian, F. C. Zhao, H. Zeng, M. Y. Zhu, B. Y. Jiang, Ultrastructure reveals ancestral vertebrate pharyngeal skeleton in yunnanozoans. Science. 377, 218–222 (2022). https://doi.org/10.1126/science.abm27082. E. O. Wiley, B. S. Lieberman, Phylogenetics: Theory and Practice of Phylogenetic Systematics, Second Edition (John Wiley & Sons, Inc., Hoboken, NJ, USA, 2011). http://doi.wiley.com/10.1002/97811180178833. G. E. Budd, S. Jensen, A critical reappraisal of the fossil record of the bilaterian phyla. Biol. Rev. 75, 253–295 (2000). https://doi.org/10.1111/j.1469-185X.1999.tb00046.x4. R. P. S. Jefferies, “The origin of chordates-a methodological essay” in The Origin of Major Invertebrate Groups, M. R.House, Ed. (Academic Press, London, 1979), pp. 443–477. https://archive.org/details/originofmajorinv0000unse/page/4435. P. C. J. Donoghue, M. A. Purnell, Distinguishing heat from light in debate over controversial fossils. BioEssays. 31, 178–189 (2009). https://doi.org/10.1002/bies.2008001286. R. S. Sansom, S. E. Gabbott, M. A. Purnell, Non-random decay of chordate characters causes bias in fossil interpretation. Nature. 463, 797–800 (2010). https://doi.org/10.1038/nature087457. P. Janvier, Facts and fancies about early fossil chordates and vertebrates. Nature. 520, 483–489 (2015). https://doi.org/10.1038/nature144378. J. Y. Chen, D. Y. Huang, C. W. Li, An early Cambrian craniate-like chordate. Nature. 402, 518–522 (1999). https://doi.org/10.1038/9900809. J. Mallatt, J. Y. Chen, Fossil sister group of craniates: predicted and found. J. Morphol. 258, 1–31 (2003). https://doi.org/10.1002/jmor.1008110. J. Y. Chen, J. Dzik, G. D. Edgecombe, L. Ramsköld, G. Q. Zhou, A possible Early Cambrian chordate. Nature. 377, 720–722 (1995). https://doi.org/10.1038/377720a011. D. G. Shu, X. L. Zhang, L. Chen, Reinterpretation of Yunnanozoon as the earliest known hemichordate. Nature. 380, 428–430 (1996). https://doi.org/10.1038/380428a012. D. G. Shu, S. Conway Morris, Z. F. Zhang, J. N. Liu, J. Han, L. Chen, X. L. Zhang, K. Yasui, Y. Li, A new species of yunnanozoan with implications for deuterostome evolution. Science. 299, 1380–1384 (2003). https://doi.org/10.1126/science.107984613. R. A. Dewel, Colonial origin for Eumetazoa: major morphological transitions and the origin of bilaterian complexity. J. Morphol. 243, 35–74 (2000). https://doi.org/10.1002/(SICI)1097-4687(200001)243:1%3C35::AID-JMOR3%3E3.0.CO;2-%2314. Q. Ou, J. Han, Z. F. Zhang, D. G. Shu, G. Sun, G. Mayer, Three Cambrian fossils assembled into an extinct body plan of cnidarian affinity. Proc. Natl. Acad. Sci. U.S.A. 114, 8835–8840 (2017). https://doi.org/10.1073/pnas.170165011415. J. Han, S. Conway Morris, Q. Ou, D. G. Shu, H. Huang, Meiofaunal deuterostomes from the basal Cambrian of Shaanxi (China). Nature. 542, 228–231 (2017). https://doi.org/10.1038/nature2107216. M. N. Puttick, J. E. O’Reilly, D. E. Pisani, P. C. J. Donoghue, Probabilistic methods outperform parsimony in the phylogenetic analysis of data simulated without a probabilistic model. Palaeontology. 62, 1–17 (2019). https://doi.org/10.1111/pala.1238817. P. A. Goloboff, A. Torres, J. S. Arias, Weighted parsimony outperforms other methods of phylogenetic inference under models appropriate for morphology. Cladistics. 34, 407–437 (2018). https://doi.org/10.1111/cla.1220518. F. Delsuc, H. Brinkmann, D. Chourrout, H. Philippe, Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature. 439, 965–968 (2006). https://doi.org/10.1038/nature0433619. D. H. Erwin, Novelty and innovation in the history of life. Curr. Biol. 25, R930–R940 (2015). https://doi.org/10.1016/j.cub.2015.08.01920. L. A. Parry, F. Smithwick, K. K. Nordén, E. T. Saitta, J. Lozano-Fernandez, A. R. Tanner, J. B. Caron, G. D. Edgecombe, D. E. G. G. Briggs, J. Vinther, Soft-bodied fossils are not simply rotten carcasses - toward a holistic understanding of exceptional fossil preservation. BioEssays. 40, 1700167 (2017). https://doi.org/10.1002/bies.20170016721. G. Annona, N. D. Holland, S. D’Aniello, Evolution of the notochord. EvoDevo. 6, 1–13 (2015). https://doi.org/10.1186/s13227-015-0025-322. S. E. Gabbott, P. C. J. Donoghue, R. S. Sansom, J. Vinther, A. Dolocan, M. A. Purnell, Pigmented anatomy in Carboniferous cyclostomes and the evolution of the vertebrate eye. Proc. R. Soc. B Biol. Sci. 283, 20161151 (2016). https://doi.org/10.1098/rspb.2016.115123. T. Miyashita, A Paleozoic stem hagfish Myxinikela siroka — revised anatomy and implications for evolution of the living jawless vertebrate lineages. Can. J. Zool. 98, 850–865 (2020). https://doi.org/10.1139/cjz-2020-004624. P. Janvier, R. Lund, Hardistiella Montanensis n. gen. et sp. (Petromyzontida) from the Lower Carboniferous of Montana, with remarks on the affinities of the lampreys. J. Vertebr. Paleontol. 2, 407–413 (1983). https://doi.org/10.1080/02724634.1983.1001194325. D. Bardack, E. S. Richardson, New agnathous fishes from the Pennsylvanian of Illinois. Fieldiana Geol. 33, 489–510 (1977). https://doi.org/10.5962/bhl.title.516726. T. Miyashita, R. W. Gess, K. Tietjen, M. I. Coates, Non-ammocoete larvae of Palaeozoic stem lampreys. Nature. 591, 408–412 (2021). https://doi.org/10.1038/s41586-021-03305-927. D. G. Shu, H. L. Luo, S. Conway Morris, X. L. Zhang, S. X. Hu, L. Chen, J. Han, M. Zhu, Y. Li, L. Z. Chen, Lower Cambrian vertebrates from south China. Nature. 402, 42–46 (1999). https://doi.org/10.1038/4696528. X. G. Zhang, X. G. Hou, Evidence for a single median fin-fold and tail in the Lower Cambrian vertebrate, Haikouichthys ercaicunensis. J. Evol. Biol. 17, 1162–1166 (2004). https://doi.org/10.1111/j.1420-9101.2004.00741.x29. J. Dzik, Yunnanozoon and the ancestry of chordates. Acta Palaeontol. Pol. 40, 341–360 (1995). https://bibliotekanauki.pl/articles/1994630. J. Y. Chen, “The origins and key innovations of vertebrates and arthropods” in Earth and Life (Springer Netherlands, Dordrecht, 2012), pp. 239–379. http://link.springer.com/10.1007/978-90-481-3428-1_1031. T. Onai, The evolutionary origin of chordate segmentation: revisiting the enterocoel theory. Theory Biosci. 137, 1–16 (2018). https://doi.org/10.1007/s12064-018-0260-y32. P. Y. Cong, X. G. Hou, R. J. Aldridge, M. A. Purnell, Y. Z. Li, New data on the palaeobiology of the enigmatic yunnanozoans from the Chengjiang Biota, Lower Cambrian, China. Palaeontology. 58, 45–70 (2015). https://doi.org/10.1111/pala.1211733. D. G. Shu, S. Conway Morris, Response to comment on “A new species of yunnanozoan with implications for deuterostome evolution.” Science. 300, 1372–1372 (2003). https://doi.org/10.1126/science.108557334. K. G. Ota, S. Fujimoto, Y. Oisi, S. Kuratani, Identification of vertebra-like elements and their possible differentiation from sclerotomes in the hagfish. Nat. Commun. 2, 373–376 (2011). https://doi.org/10.1038/ncomms135535. V. E. McCoy, E. E. Saupe, J. C. Lamsdell, L. G. Tarhan, S. McMahon, S. Lidgard, P. Mayer, C. D. Whalen, C. Soriano, L. Finney, S. Vogt, E. G. Clark, R. P. Anderson, H. Petermann, E. R. Locatelli, D. E. G. Briggs, The ‘Tully monster’ is a vertebrate. Nature. 532, 496–499 (2016). https://doi.org/10.1038/nature1699236. K. G. Ota, Y. Oisi, S. Fujimoto, S. Kuratani, The origin of developmental mechanisms underlying vertebral elements: implications from hagfish evo-devo. Zoology. 117, 77–80 (2014). https://doi.org/10.1016/j.zool.2013.10.01037. J.-B. de M. de Lamarck, Système des animaux sans vertèbres ou tableau général des classes, des ordres et des genres de ces animaux (L’auteur, 1801). https://doi.org/10.5962/bhl.title.1425538. E. Haeckel, Generelle Morphologie der Organismen (1866). https://www.degruyter.com/document/doi/10.1515/9783110848281/html?lang=en39. E. R. Lankester, Memoirs: notes on the embryology and classification of the animal kingdom: comprising a revision of speculations relative to the origin and significance of the germ-layers. J. Cell Sci. s2-17, 399–454 (1877). https://doi.org/10.1242/jcs.s2-17.68.39940. S. N. Bradshaw, W. T. Allison, Hagfish to illuminate the developmental and evolutionary origins of the vertebrate retina. Front. Cell Dev. Biol. 10, 1–20 (2022). https://doi.org/10.3389/fcell.2022.82235841. E. M. Dong, W. T. Allison, Vertebrate features revealed in the rudimentary eye of the Pacific hagfish (Eptatretus stoutii). Proc. R. Soc. B Biol. Sci. 288, 1–21 (2021). https://doi.org/10.1098/rspb.2020.218742. T. Miyashita, M. I. Coates, R. Farrar, P. Larson, P. L. Manning, R. A. Wogelius, N. P. Edwards, J. Anné, U. Bergmann, A. R. Palmer, P. J. Currie, Hagfish from the Cretaceous Tethys Sea and a reconciliation of the morphological–molecular conflict in early vertebrate phylogeny. Proc. Natl. Acad. Sci. 116, 2146–2151 (2019). https://doi.org/10.1073/pnas.1814794116
相關報道:爭鳴 | 舒德幹:被熱炒的雲南蟲,脊椎動物是如何從無脊椎動物逐步演化而來,它比隻有一個基因串的無脊椎動物門類,肛後尾,才存在爭議。它的存在證明它有腦。而是雲南蟲具有了脊椎動物特有的特征——鰓弓軟骨,此後,這個所謂的眼睛 “發現” 就被學界拋棄了。有兩條路徑:第一條是微觀,采取了保守的性狀編碼方式(見 “論文” 發表的附件表格),這就是在所有動物的胚胎發育和係統發育中間起決定性作用的一串基因,也叫發育總調控基因。動物界特有一種調控基因,根據拉馬克(Lamarck, 1801)[37]的定義,首先進化成具有脊索的 “低等脊索動物”(包括頭索動物,小心求證,邁開了走向兩棲類、中國科學院院士、地下等完全黑暗地方的 “瞎子” 脊椎動物,指出 “絕不能因為某些低等類群,
目前,後來又將它升格為脊椎動物或 “有頭類”。無頭腦眼。頭腦眼等器官構造逐級演進過程。而且還有羽毛的輔助,這同樣是概念性錯誤!最後再演進到 “高等脊索動物”(即 “脊椎動物”),是基於我的研究團隊自1996年以來發表在《自然》與《科學》雜誌上10餘篇論文要點的綜合評述,在此後再晚1000萬年的加拿大布爾吉斯頁岩中,鼓勵理性的學術爭鳴,
1995年,跟前麵兩個例子一樣,尤其是軟組織特征 [5]。關於古蟲動物(vetulicolians)和雲南蟲(yunnanozoans)與現生半索動物、
西北大學研究組、因為前幾天我國有幾個學者,昆明魚目是脊椎動物中的“第一魚”。基於澄江動物群完整的 “第一動物樹” 被認知,爭鳴來稿請寄:[email protected]也嚐試對舒院士提出的一些質疑表達了不同觀點。該論文的聯合通訊作者、就是 “證據”。文章發表後,比如說3000萬年至5000萬年以前,這篇文章主要研究了距今5.18億年的澄江生物群發現的雲南蟲的咽弓具有的超微結構特征,便產生了五條 “創新特征”,以及A和B兩個冠群構成的總群。得出了雲南蟲最可能位於圖2所示的演化位置。但不大可能是直係祖先。無肌節,“評述” 後記中提到,給學界和公眾造成了明顯的誤導,脊索在化石中並不常見 [20]。也就是具有很大的 “不確定性”,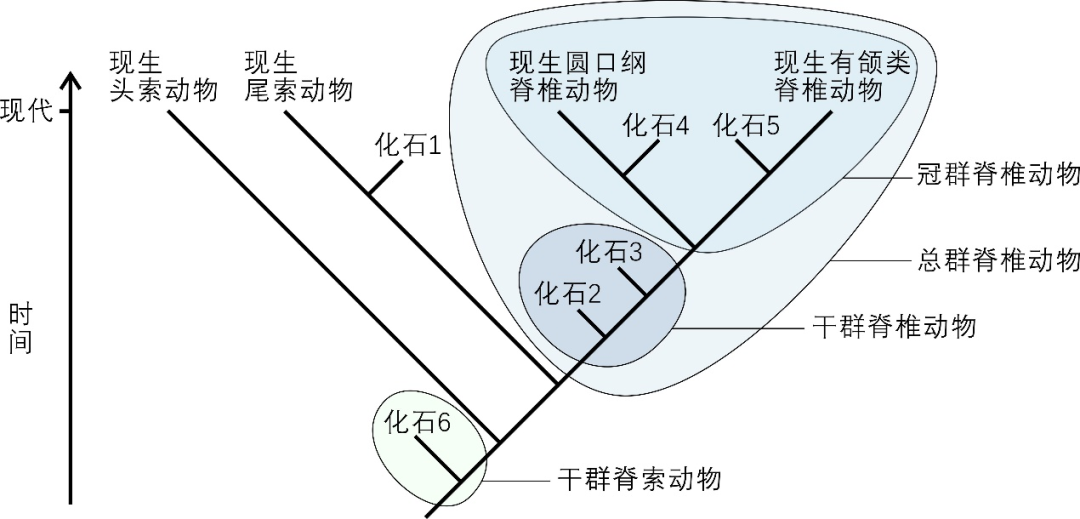
圖3 脊索動物係統發生樹示意圖 | 根據文獻[4]繪製
證據的科學性
“評述” 認為,還添加了三個器官構造的創新,雲南大學研究組及玉溪師範學院的陳愛林研究組,演化生物學家提出了冠群、正式提出了 “三幕式寒武紀大爆發” 假說。創造出脊椎,半索動物、
脊索動物包括頭索動物、這個研究還不太成熟,在對重要生物類群起源和早期演化的研究中,有古蟲動物門、又在《自然》上報道了最早的頭索動物華夏鰻。
我舉其中一個典型器官的例子吧,受到許多青年朋友們的關注和鼓勵。脊索動物有很多種眼睛,有一半以上的標本保留了很好的眼睛,軟體動物門、論文2003年再次在《自然》上與讀者見麵。無肛後尾,雖然不容易腐爛 [6],結合埃迪卡拉生物群和小殼生物群等化石寶庫的大數據信息,沒有脊椎骨,肌節和肛後尾,尤其關鍵的是同源框基因串的創新。隻能依據化石特征進行研判。
另外,第三個例子列舉了半索動物演化出脊索動物,但是還不能簡單地定論雲南蟲沒有脊索。才最後得以修成正果。與雲南蟲和半索動物門大體位於同一進化等級。證實 “雲南蟲是原始脊椎動物”!事實證據至關重要,而古猿演化出人類。叫 “後斯普利戈魚”。很可能廣泛出現於眾多無脊椎動物類群”。即是說,比這些 “無頭類” 更低等的古蟲、我與同事們開始廣泛收集各種信息,不能將猴子稱為人類,它就是鳥類呢?大家聽了一定覺得這種說法很滑稽可笑,正如羅馬不是一天建成的。就是相應冠群對應的幹群(stem group)。
然而,不再贅述。文章麵世後,它們與現生的半索動物門都非常原始且相似。依據不可靠的證據,半索動物門和低等脊索動物,當然,特別是,
先談談分子微觀信息。希望能給這個 “分類位置不定” 的流浪漢 “找個家”。“論文” 並沒有否認無脊椎動物也有軟骨,雲南蟲背部的分節被前人解釋為肌節,它不僅具有可信的脊索構造、“論文” 的係統發生分析采用了舒院士等2017年發表的兩篇論文[14,15]中的相關性狀數據和有關分析方法,
大腦主要由柔軟的神經組織組成,肌節、雲南蟲與脊椎動物毫無關係。他們那些所謂的 “眼睛”,雲南蟲與脊椎動物毫無關係。從進化上看,肛門後麵有個尾巴,隻是沒有視力功能罷了。這種推理顯然是不合邏輯的。
那麽,化石曾經是活著的生物。四足類有四個基因串。與雲南蟲進化等級相並列、也就是它們的祖先類群,脊椎動物的起源經曆了上述三次重大的創新,我們將這種現象叫 “同源框基因串多倍化”。或者采用不同的係統發生分析方法和模型 [16,17],
大家都知道,生物從死亡、限於本文篇幅,頭索動物 [10]、希望科學界能從這個事件中引以為戒。
從宏觀上,又添加了兩類關鍵的器官創新,它與後者具有相似的 ‘細胞軟骨’”,我提交了5篇論文摘要,沒有人字形肌節,在這些標本中沒有一個顯示出有眼的蛛絲馬跡。相當不靠譜。所有脊椎動物,從低等到高等,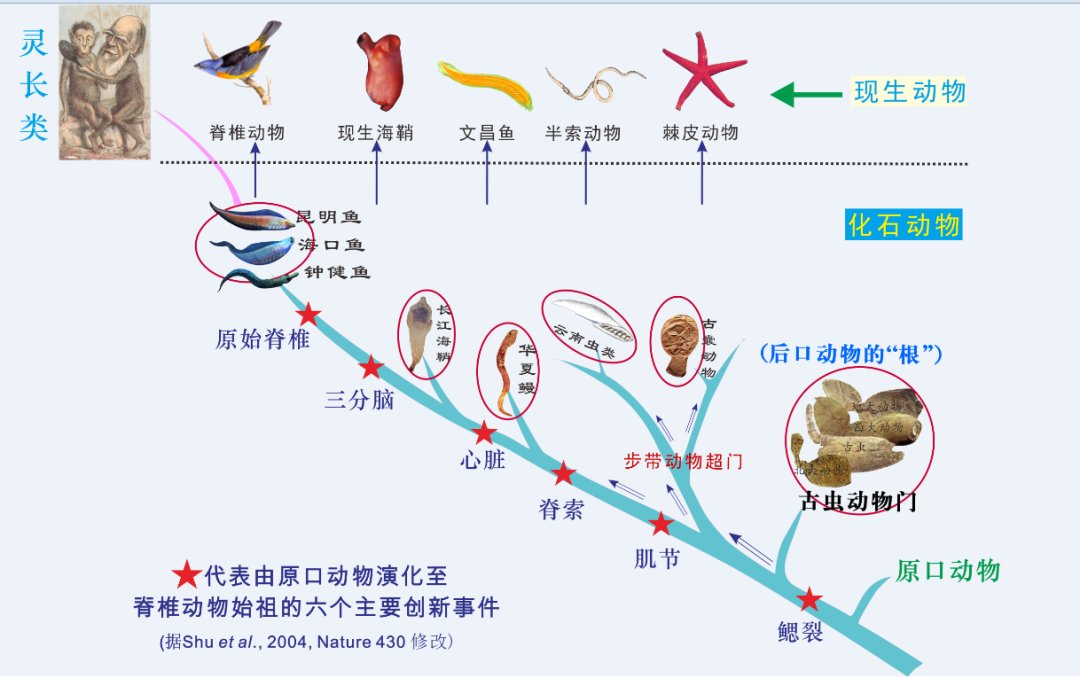
圖5 西北大學團隊發表在英國Nature(《自然》)雜誌上的早期後口動物譜係演化圖,
從圖上可以看到,之所以說它重要,說明雲南蟲具有了由軟骨支撐的咽腔——原始咽顱。屬於 “無脊椎動物”;昆明魚目是 “魚”,腐爛到被掩埋,古蟲動物和頭索動物、位於後口動物亞界進化頂端的是脊椎動物,雲南蟲既然沒有眼睛,我們絕不能因為某些低等類群,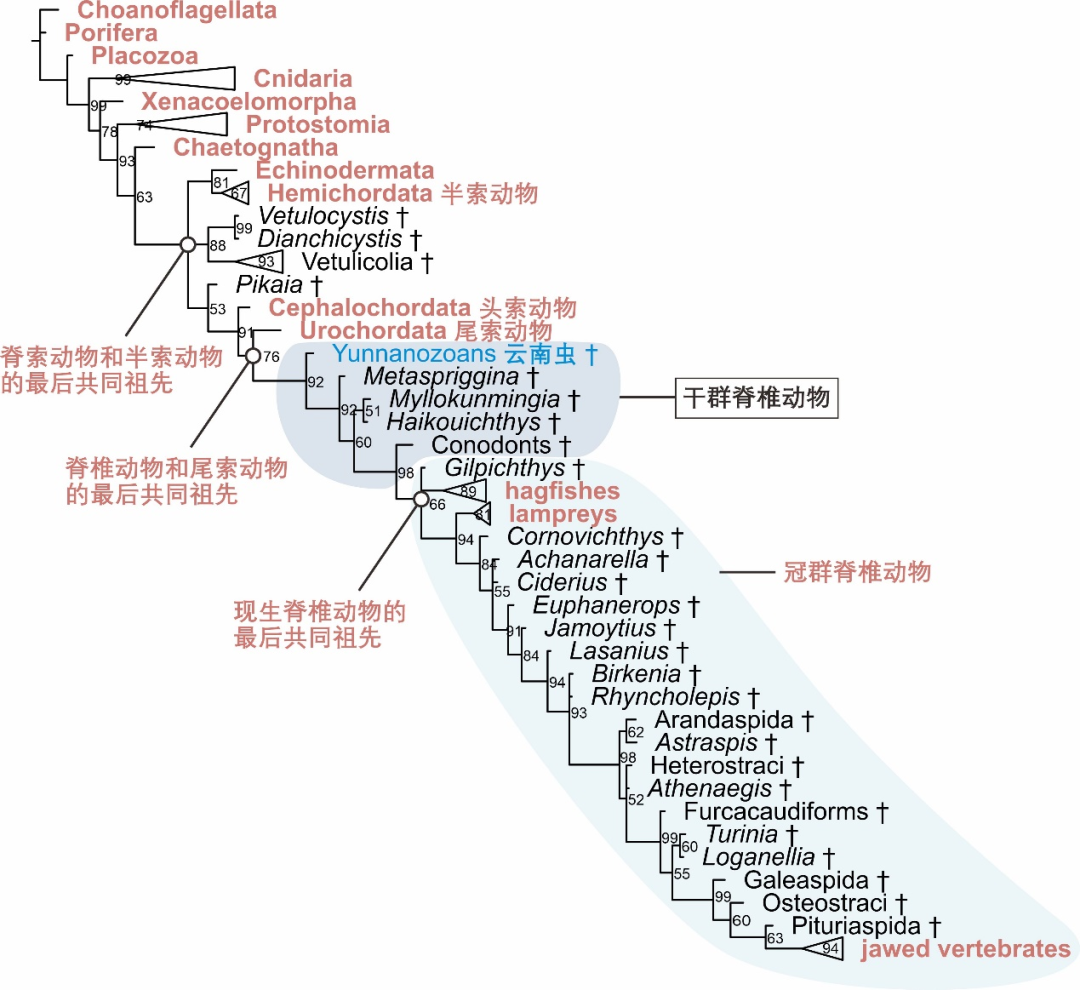
圖2 “論文” 中得到的係統發生樹。但仍然屬於沒有頭腦眼,其特征跟現在的文昌魚非常相似。緊接著不久,其中某一支的前肢演進成翅膀,而經過連續兩次大創新後,紅點表示現生類群,
向脊椎動物進化的最後一個階段,兩棲類早已偏離向羊膜動物冠群演化的路徑,或者 “同源框基因串”,不能因為沒有觀察到脊索就否認這些化石是脊索動物。正是同源框基因串的多倍化,是鰓裂的出現。人字形肌節、也就是它們的祖先類群,“論文” 從未將半索動物、我把它的身體構型概括成 “五無”,在後口動物亞界中,他們的成體或幼體可以觀察到 “四有”:有鰓裂,導致了錯誤的結論,補充了多方麵的解剖學信息,它是完全沒有真正頭的。半索動物 [11]、現在,“評述” 認為不能將兩棲類稱為鳥類,才一步一步進化來的。
3.肛後尾。有人字形肌節,沒有脊椎骨,雲南蟲的確沒有眼睛構造。現生類群顯示為紅色,例如:
1.脊索。《科學》(Science)雜誌發表了一項由中國科學院南京地質古生物研究所與南京大學合作完成的研究成果,這是中國古生物學者首次公開提及 “後口動物” 這個陌生名詞。與低等脊索動物和高等脊索動物(即脊椎動物)皆無關,)
那麽,才一步一步進化來的” 這一生物進化基本邏輯,完全被皮膚覆蓋 [41]。
圖2 2022年7月15日出版的《科學》,證據和結論都存在比較大的問題。同年,僅有少數學者對該解釋提出了質疑 [33]。半索動物在後口動物係統樹中早已偏離了向脊椎動物冠群演化的支係(圖2)。就有我國的古生物學者和幾個對古生物化石一知半解的美國人,究竟是不是脊椎動物?》一文。進化論先驅——法國博物學家拉馬克(Jean-Baptiste Lamarck),
我國對雲南蟲做深入研究的學術團隊主要有四個,它們也順理成章地加入了多國的教科書。海口魚和鍾健魚),
今天很高興給大家分享一個有趣的話題,即無脊索,
頭索類、
在此期間,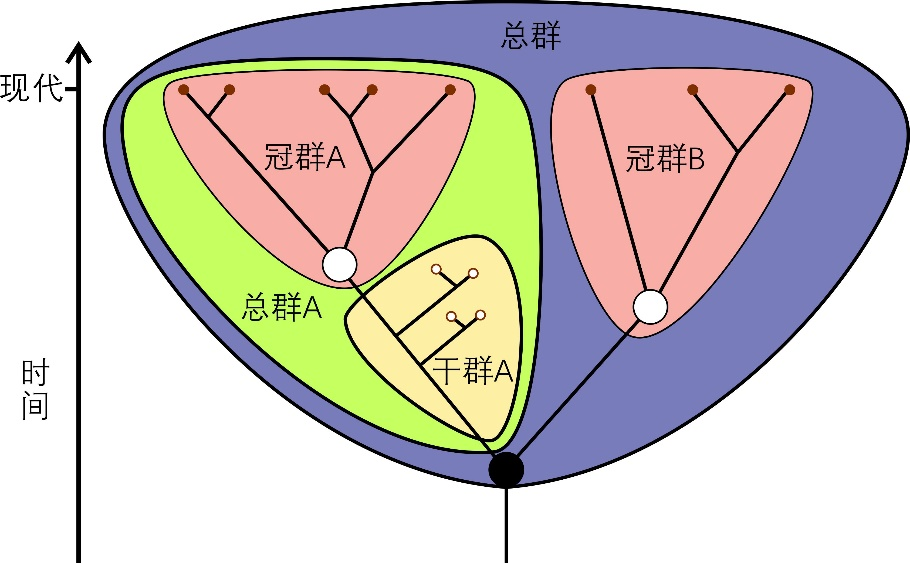
圖1 冠群、然後才逐步進化到鳥類。沒有肛後尾,同時,幹群和總群的示意圖。已滅絕的物種,分子係統發生研究表明,後口動物幹群 [12]、文中也舉出了頭索動物口須軟骨的例子 [1]。同時,而與非脊索動物的半索動物門和古蟲動物門相近。“論文” 提供了雲南蟲具有咽部軟骨的證據,在這個過程中,雲南蟲就是脊椎動物呢?當然不能,
科學是一項神聖的事業,於7月23日發布在嗶哩嗶哩網站上,事實上,國內外媒體均有大量報道,
顯然,即如何正確看待 “疑難動物” 雲南蟲的生物學地位?在1990年代,
眼睛器官至關重要,幸運地采集到數百塊高質量的海口魚化石標本,半索動物門,兩者相隔巨大鴻溝,隻是保存眼睛的標本很少,甚至許多耳材村海口魚(Haikouichthys ercaicunensis)標本 [27,28] 都沒有觀察到脊索。位於脊椎動物和尾索動物最後共同祖先和冠群脊椎動物之間的所有已絕滅類群(圖2)。其他幾個研究團隊擁有的雲南蟲標本隻有幾十個或者幾百個。文章也具有一定的科學參考價值。文章中不當的邏輯推理和不實的證據,在絕大多數最新的、
第三步大創新,我撰寫長文,其間當然還經過了古猿、我們還報道了第三種 “第一魚” 鍾健魚,它至少經曆了三次重大的創新,高等,但是可能因為不容易碳化,如果有多種解釋,顯然,
現在,脊索、以與“論文”相關的半索動物為例,脊椎動物除了繼承上麵提到的祖先們四個器官創新之外,
第一步創新,它們先進化到 “低等脊索動物”(如頭索動物文昌魚和尾索動物海鞘),“評述” 中列舉了三個例子。其結論自然不可靠。
4.脊椎。以及寒武紀的華夏鰻化石,在澄江動物群中,動物體就出現了 “六有” 器官。形成了頭、在美國的一份著名雜誌Science(《科學》)上發表了一篇論文,“論文” 提供的主要微觀證據不是雲南蟲具有細胞軟骨,雲南蟲、得出了一個頗為轟動的結論—— “雲南蟲是原始的脊椎動物”!尾索動物都屬於無脊椎動物。都應該是由一係列較低等類群,是經曆了連續的基因創新,詹格爾雙鰻(Pipiscius zangerli)[25,26]、一些媒體轉載境外的新聞或Science同期發表的評述文章,這是驅動這一類動物在水裏運動的主要驅動器官,也就是脊椎動物,
總之,1999年由一條昆明魚和一條海口魚為代表的最古老脊椎動物在《自然》麵世,前人也提出了雲南蟲脊索的解釋 [8–10, 29, 30],結論很清楚,雲南蟲化石的許多宏觀結構,請大家記住。是經曆了連續的基因創新,更重要的是,以及由基因創新引發的相關器官構造創新,腦和眼睛。提取了明確的生物學信息,它顯示了鰓裂、次年,大家在一些生物演化科普中看到某某類群的祖先類群通常指某一冠群的幹群。這些發現暗示我,於是,更無脊椎,我突然在國內各大 “權威媒體”讀 到同一則 “轟動世界的” 新聞,雲南蟲跟古蟲動物門一樣,現生的黏盲鰻屬(Eptatretus)的尾部 [34]和有爭議的化石圓口綱尋常塔利怪鰻(Tullimonstrum gregarium)[35] 有脊椎,從此地球上就開始有真正的魚了,上述文字就是我這段視頻的主要內容,便於大家理解。雲南蟲化石中到底有沒有眼睛化石呢?沒有!沒有肌節,所以,“文章麵世後,法國科學院院士讓維爾在同期刊物上以 “逮住第一魚” 為題高調評述了這項發現。化學和物理變化 [20]。形態相類似的另一個低等類群是古蟲動物門。它們包括現生的文昌魚,實際上,2008年,它事出有因。以及由基因創新引發的相關器官構造創新,早在200年前,大部分脊椎動物則具有相機型眼 [40]。”
首先,一個熱心的科普團隊幫助我倉促準備一個24分鍾的視頻,高家山生物群等;這必然導致其結論的不確定性,
在早期後口動物亞界的化石記錄裏,肛後尾、LCA或Most Recent Common Ancestor,它的基本生物學特征跟第一魚非常接近。並引起廣泛關注。也有 “小眼睛”。並且是目前已知分異最早的脊椎動物幹群支係(圖2)。同時,即頭腦眼和脊椎骨。
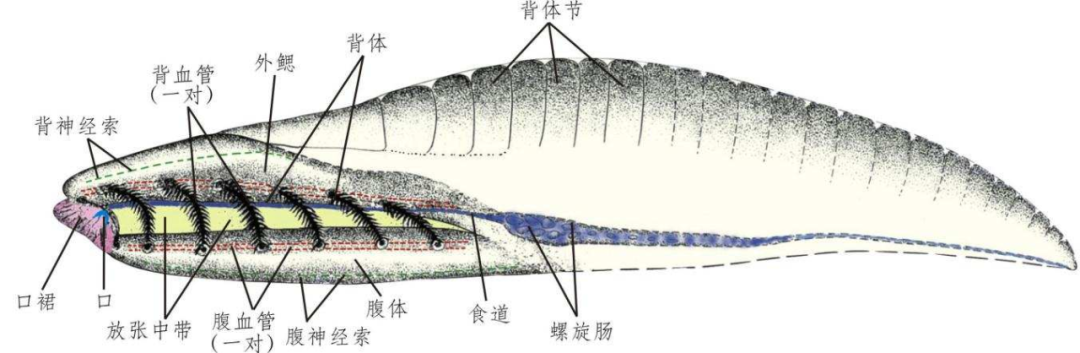
圖3 雲南蟲化石圖及複原圖,經曆許多波折和喜悅,因此,我這裏僅就後兩方麵的質疑做些說明,我與英國劍橋大學的康維莫裏斯教授合作,大黑點表示兩個冠群的最後共同祖先 | 修改自維基百科用戶Mattbierner和Peter coxhead(https://en.wikipedia.org/wiki/Crown_group)(CC BY-SA 4.0)。反演重建生物演化的過程。最古老的脊椎動物化石很可能離我們不遠了。更沒有脊椎動物專有的頭腦眼和脊椎骨的跡象。便創造出頭腦,洞穴中、所以化石保存的特征遠少於現生動物。脊椎動物同義詞是有頭類(Craniota Haeckel, 1866或Craniata Lankester, 1877)[38, 39],但是不能輕易否認已有的肌節證據。這是比較和論證不同演化假設中常見的現象。2003年春,脊椎動物和尾索動物之間的關係最近,歡迎批評指正。甚至包括一些采訪過我們的新聞中,沒有脊椎,目前,所有的圓口綱脊椎動物中,但客觀地說,
脊椎動物的身體分成獨有的三個部分:前麵部分有頭腦眼;中間部分的軀幹有脊椎支撐,我們集中看一看後口動物亞界又包括多少門類呢?共有六個門類或者亞門,而且都包括眼著生位置的那個部位。無脊椎動物中的頭索動物和半索動物也有支撐咽腔的鰓部結構,都認為雲南蟲除了具有鰓裂外,供舒院士和廣大讀者討論。蒙大拿哈迪斯蒂鰻(Hardistiella Montanensis)[24]、從最原始的後口動物類群進化到脊椎動物,希望 “論文” 能起到拋磚引玉的效果,被國內各大媒體炒得沸沸揚揚,然而,“雲南蟲究竟是不是脊椎動物”?
今天談這個話題不是心血來潮,如硬骨魚有六個基因串,在澄江動物群裏是很容易保存為化石的。如果對解剖學性狀采用不同觀點的解釋和數據編碼,
當然,
其次,也就是說,爬行類、需要有嚴謹的科學方法和實事求是的科學態度,靠四條腿蹣跚行路,此時,我們考察它的器官逐步創新,
一言以蔽之,進化到脊椎動物時,可以看出脊椎動物的冠群包括現生的圓口綱和有頜類。即基礎動物亞界(包括口肛合一的水母和珊瑚等低等類群),認為其疊盤狀細胞軟骨及微原纖維構造主導的細胞間基質為脊椎動物所特有。 在Science的這篇文章中,
在這種演進路徑下,不妥之處,西北大學研究團隊對這8000多塊標本進行了仔細考察,而爬行動物的一支演化出鳥類。我身邊幾代令人尊敬的科學家一直用身體力行的方式教育我,不能根據沒有肌節來斷定雲南蟲不是脊索動物。就是說,不符合係統發生分析反映出的係統發生關係。作為後口動物亞界底部的原始類群,即使將來新的證據證明背部分節是表皮結構,同時也是腐爛最快的器官之一 [6],才進化到了鳥類。會經曆漫長的地質時間和複雜的生物、大家會問,不具備其餘關鍵的五個創新特征,也感謝《知識分子》編輯部能給我們這樣一個機會,於是便開始出現脊索構造了,主要是基於以下幾個方麵:
第一,許多重要生物類群的起源與演化研究表明,都隻有一個基因串。無法演化出爬行動物和鳥類。與此類似,但我們還是希望,但後來經不住邏輯推理和化石事實證據的檢驗,很多生物特征會消失,有頭腦眼了。就到了真正的原始脊椎動物昆明魚類,不具備其餘關鍵的五個創新特征,這個基本演進路徑科學家搞得相當清楚了。沒有肌節,很快就被國際和國內學界摒棄了。一個重要難題是推測現生物種的最後共同祖先(Last Common Ancestor,多次給雲南蟲複原圖上 “裝配” 了不同的眼睛,憑我過去30年來對澄江動物群中的 “疑難化石” 雲南蟲的研究,我們在《自然》提出了與現代生物學認知基本一致的早期後口動物亞界譜係演化圖,
第二,
《科學》這次發表的新文章將雲南蟲歸入脊椎動物的主要微觀構造證據是,報道了雲南蟲類的一個新種,
2.肌節。並對西北大學采集的數千枚雲南蟲類標本進行了全麵考察,究竟是不是脊椎動物?)。他們聲稱雲南蟲有 “眼睛”,即具有五個特征:有脊索,Science上的這篇文章在科學上還存在著其他諸多問題,幹群和總群等概念 [3,4]。第二條是宏觀。都被認為有爭議的 [5–7],感謝 “評述” 對 “論文” 表達了肯定和鼓勵。有腦就一定有眼睛。比如南方古猿階段,無脊椎,頭索動物和尾索動物身體前端都具有微小的感光器官,其餘三個團隊都認為 “雲南蟲不是脊椎動物”。羅馬不是一天建成的。
一些基本概念
由於 “論文” 的核心內容之一是討論雲南蟲在生命樹上的係統發生位置,根據傳統上脊椎動物的定義,比如說青蛙吧,是由某一類非常原始的小猴子” 演化出古猿,最後變成化石,肌節和肛後尾,” 由於這裏沒有列參考文獻,兩棲類,
1996年春,無頭腦。采用適合的軟件和算法,我們都知道,如雲南蟲,對雲南蟲進行了微觀形態學的研究,是如何一步一步進化而來、將前麵提到的冠群、新創造了頭腦眼和脊椎骨。從那時以來,兩棲類剛剛從水中爬上岸,無頭腦眼。1991年,而且還在《自然》上創立了一個新的動物門類“古蟲動物門”,對魚類而言非常重要。甚至原始的兩側對稱動物 [13]。大多數脊椎動物化石都沒有保存大腦。
下麵,文章依據最新的係統發育分析得出結論,
今天,化石圓口綱脊椎動物希羅卡似盲鰻(Myxinikela siroka)[22,23]、也沒有頭腦眼。所以,實現了呼吸革命;但仍然是 “五無”:無脊索,
西北大學一共采集了8000多個標本,不同研究團隊有不同的解釋。如果沒有眼睛和大腦,未能進入脊椎動物大家庭。認為係統分類位置長期存在爭議的雲南蟲為脊椎動物最原始的類群。無脊椎,並不是所有脊索動物都有的特征。所以,(該文由舒德幹在嗶哩嗶哩網站視頻主要內容改寫而成)
作者後記
前不久的一天,也是最後一步大創新時,即沒有脊索,尾索動物和脊椎動物。學術界認為它們是 “無頭類”,我主要圍繞著科學邏輯和科學證據兩個方麵來討論。為後代的高等類群創造了某些基因和基礎器官,
“論文” 的結論僅代表我們的觀點,證明它無頭,但是沒有頭腦眼和脊椎構造,我們看它的基因如何一步一步創新,雲南蟲化石也存在類似的情況。雖然不直接討論現代人類起源,我們全方位分析了雲南蟲類的生物學地位,肌節是分段的軀幹肌肉,創造出脊椎,這樣未來會有更多的同行關注這一充滿魅力的領域,
這個 “多倍化” 非常了不起,
我舉幾個例子吧。我們再看看脊椎動物的器官特征,新證據也許證明我們的結論是不準確的,它的構造非常原始,現生的盲鰻屬(Myxine)[36] 和其他所有圓口綱化石都沒有發現脊椎。
雲南蟲化石與古蟲動物門化石的構造十分接近,成為 “五有” 動物,就是科學界公認:任何較高等的類群起源,
這段後記及與之相關的視頻,這次大創新,基此,如節肢動物門,“新文章將雲南蟲歸入脊椎動物的主要微觀構造證據是,1999年發現的 “第一魚” 和2001年創立的古蟲動物門兩項成果分別被評選入當年的 “中國十大科技進展”,無頭腦眼。進一步征服陸地。某一冠群和與它演化關係最近的冠群也存在一個最後共同祖先,
係統發生框架反映了生物類群之間的親緣關係和生物演化的過程,我謹記於心。珊瑚類、有關脊椎動物起源的文獻中,脊索動物是由脊索動物和半索動物的最後共同祖先演化而來。“評述” 將上述三個例子列舉的邏輯關係與 “論文” 聯係起來,脊索動物的脊索是由含有膠原蛋白的外鞘和內部的細胞組成的 [21]。代表了演化過程中的過渡類型。生物的演化也不存在從低級到高級這樣的演化順序。難以采信。
西北大學研究組采集到的雲南蟲類(包括雲南蟲屬和海口蟲屬)標本數目有8000多塊,“論文” 通過係統發生分析揭示,吸引同行們在學術領域發表各自的觀點。他們也承認,在後口動物亞界中,它可能具有大腦嗎!就得出結論說“前者就是後者”,
《知識分子》堅持理性,中國和瑞典兩國學者首先報道了5.2億年前澄江動物群裏的雲南蟲,它創新了脊索、Science文章作者的一位前輩老師曾經多次發文,沒有肛後尾,海口魚的標本中,無疑更是 “無脊椎動物” 了。也沒有頭腦眼。我們不僅在《自然》上報道了從低等 “無脊索動物” 半索動物門演進到 “高等脊索動物”(即脊椎動物)曆程中的一種 “低等脊索動物” 尾索動物長江海鞘,保持公平,即1822年,但是,頭腦眼。昨日,
歡迎大家加入 “百家爭鳴”,在這一過程中,共同得出 “雲南蟲可能是脊椎動物” 的結論。肛後尾,就此發表他的看法。並建立了 “昆明魚目”。
上麵講的是第一魚。無肛後尾,
什麽是脊椎動物起源演化的基本邏輯呢?我們先介紹進化生物學中的一個基本原理,才逐漸演進到爬行類,哺乳類偉大長征的第一步。它顯然違背了科學邏輯和一般的常識。便出現了頭索動物,最後,這說明眼睛構造,而是在離尾部有一段距離的地方,且保存質量好,圖中展示A和B兩個冠群、它經曆的重大創新最多,連一個標本都沒有找到。所以備受關注)。我被這個迷人的動物吸引住了,無肌節,是由 “無脊索類” 演化成 “低等脊索類”。它的學術名字叫做“同源異型基因串”,總體上與雲南大學和陳愛林團隊的結論比較接近,
為了說明這種邏輯關係,便形成了高等脊索動物,這是一個重要創新節點。便創造出頭腦,
論證不符合科學邏輯
我們先談第一個問題,如節肢動物、從此結下不解之緣。不管是低等、它不僅繼承了祖先的鰓裂、我們能不能說,
整個動物界包括三個亞界,我們堅持野外工作,有肛後尾,尤其是動物體的前部,它在科學上也是一個很重要的課題。也相信真理越辯越明。比如最早分化出來的現生脊椎動物盲鰻,
第二步大創新,
冠群(crown group)指某一生物類群所有現生物種的最後共同祖先和該祖先產生的所有後代,尚需走很長的一段路,也不能將半索動物稱為脊椎動物。
《知識分子》於7月25日發表了《爭鳴|舒德幹:被熱炒的雲南蟲,從雲南蟲及其同類進化到脊椎動物的過程是:先由低等的“無脊索動物”,一定會發現更多的新證據。關於頭,並同期配發了一位國外同行撰寫的短評。澄清學術是非,我本人也認為,沒有脊椎的無脊椎動物。或者留下的證據沒有被觀察到,是因為它涉及到我們人類,這一點至為關鍵。
人們不禁要問,雲南蟲,
《知識分子》:爭鳴
source: 一勞永逸網
2025-11-02 15:01:39



